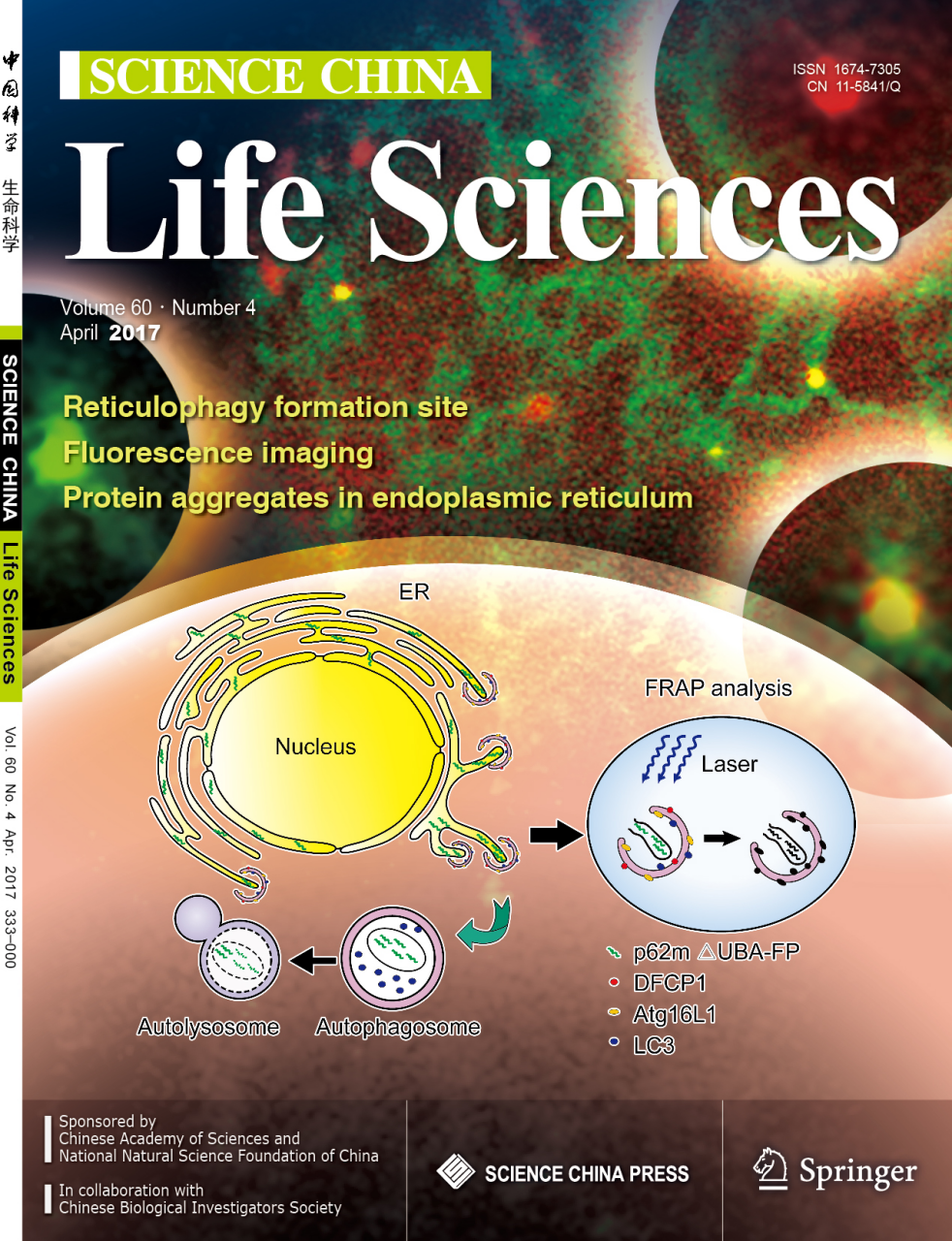
华中科技大学武汉光电国家实验室(筹)生物医学光子学功能实验室张智红教授课题组,建立了一种蛋白质诱导的内质网自噬模型,并在活细胞内通过荧光成像方法揭示了内质网自噬的发生地点,为“内质网作为内质网自噬的发生地点”这一观点提供了实验依据。论文“Visualization of reticulophagy in living cells using an endoplasmic reticulum-targeted p62 mutant”于2017年4月1日作为封面文章发表在SCIENCE CHINA Life Sciences杂志上(60: 333–344, 2017)。
自噬作为真核细胞降解细胞内蛋白和膜系统等物质的生理性过程,对于维持胞浆中蛋白和细胞器的新老平衡及在低营养条件下提供基本营养分子(如氨基酸和磷脂分子等)有着重要的作用。自噬过程的本质是在细胞质中形成自噬膜来包裹胞浆中的蛋白质或者受损的细胞器,然后将其运输到溶酶体中进行降解。现有的研究表明,几乎任何一种细胞器都可以被自噬囊泡包裹。其中,关于一些重要的细胞器如内质网或者线粒体等的自噬降解得到了广泛的关注。本研究中,我们重点关注内质网自噬及其发生地点。
在内质网受压条件下,堆积了过多的错误折叠的蛋白质或者蛋白聚集体时,内质网的自我保护机制被启动。例如,当未折叠蛋白反应(UPR)和内质网相关的降解(ERAD)信号通路处于饱和时,内质网自噬便成为降解这些功能缺陷的内质网的主要途径。已知一些化学药物和突变蛋白质的表达能够引起内质网中蛋白聚集体的形成并诱导自噬过程。但是,关于内质网自噬的发生地点仍然不很清楚,一般认为有两种可能,一种是自噬膜在内质网膜处生成并包裹后者,另一种是受损的内质网从内质网系统上剥离出来并被运输到自噬膜处。针对上述问题,张智红教授及其博士研究生王亮等人,利用自噬接体分子(p62/SQSTM1)的寡聚化性质,获得了一个突变体p62m∆UBA-FP,该突变体在细胞中表达后定位于内质网并且保留了寡聚化的能力。由此,我们建立了一种荧光蛋白标记的内质网自噬模型,用于活细胞内可视化研究与确证自噬的内质网起源及其命运。研究结果发现,p62m∆UBA-FP在内质网腔中形成寡集体,它不仅与内质网的标志蛋白mLumin-KDEL/mCherry-sec61 beta共定位,还与自噬的早期标志分子(如DFCP1,Atg16L1)和中晚期标志分子(如LC3)共定位。最终,绝大部分p62m∆UBA-FP被运输到了溶酶体,进一步被降解和清除。活细胞动态荧光成像研究表明,p62m∆UBA-FP寡集体及与其共定位的自噬蛋白,均与内质网结构有着紧密的物理联系,这意味着自噬膜在内质网处形成并包裹将要被降解的内质网局部,为 “内质网作为内质网自噬的发生地点”这一观点提供了实验依据。
该研究结果作为封面文章发表在中国科学-生命科学杂志上,研究工作得到了国家自然科学基金重大研究计划(91442201)、国家杰出青年科学基金(81625012)和国家自然科学基金委员会创新研究群体科学基金(61421064)等项目的资助。